(витамин РР, от англ. pellagra preventing -предупреждающий пеллагру), группа биологически активных соед., включающая никотиновую кислоту и ее производные. Наиб. распространены в природе сама к-та и ее амид, обладающие одинаковой витаминной активностью.
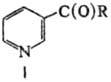
Н и к о т и н о в а я к-та (мол. м. 123,11; ф-ла I, R = OH)-бесцв. кристаллич. в-во, т. пл. 236-237 °С. Водный р-р имеет максимумы поглощения при 261,5 нм (e = 3,1Х103 при рН 5,6) и 385 нм. Н и к о т и н а м и д (мол. м. 122,13; I, R = NН 2)- бесцв. кристаллич. в-во, т. пл. 128-131 °С; хорошо раств. в воде и этаноле, плохо-в хлороформе, ацетоне и эфире. Для водного р-ра максимумы поглощения при 261,5 нм (e = 2,85.103 при рН 5,72) и 300 нм. О
…
Далее
(витамин РР, от англ. pellagra preventing -предупреждающий пеллагру), группа биологически активных соед., включающая никотиновую кислоту и ее производные. Наиб. распространены в природе сама к-та и ее амид, обладающие одинаковой витаминной активностью.
Н и к о т и н о в а я к-та (мол. м. 123,11; ф-ла I, R = OH)-бесцв. кристаллич. в-во, т. пл. 236-237 °С. Водный р-р имеет максимумы поглощения при 261,5 нм (e = 3,1Х103 при рН 5,6) и 385 нм. Н и к о т и н а м и д (мол. м. 122,13; I, R = NН 2)- бесцв. кристаллич. в-во, т. пл. 128-131 °С; хорошо раств. в воде и этаноле, плохо-в хлороформе, ацетоне и эфире. Для водного р-ра максимумы поглощения при 261,5 нм (e = 2,85.103 при рН 5,72) и 300 нм. Оба соед. весьма устойчивы при хранении и выдерживают нагревание в водных р-рах при 120°С. В р-рах к-т и щелочей никотинамид превращ. в никотиновую к-ту.
Биол. роль никотиновой к-ты и никотинамида обусловлена их участием в биосинтезе никотинамидных кофермен-тов (коферментных форм Н.): н и к о т и н а м и д а д е н и н-н у к л е о т и д а (НАД; II, R = Н) и н и к о т и н а м и д а д е н и н д и н у к л е о т и д ф о с ф а т а [HАДФ; II, R = PO(OH)2].
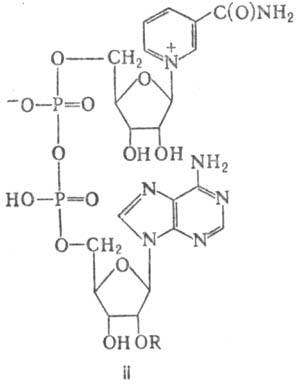
НАД и НАДФ - коферменты многочисленных (более 100) де-гидрогеназ, функционирующих на начальных этапах биол. окисления самых разнообразных соед.: углеводов, аминокислот, жирных к-т и др. Среди них ферменты гликолиза ( глицеральдегидфосфатдегидрогеназа, лактатдегидрогеназа), пентозофосфатного цикла окисления углеводов (глюкозо6-фосфат-дегидрогеназа, 6-фосфоглюконатдег идрогеназа), цикла трикарбоновых к-т (малат- и изоцитратдегидрогеназы) и др.
В катализируемых этими ферментами окислит.-восстано-вит. р-циях НАД и НАДФ играют роль промежут. акцепторов и переносчиков электронов и атомов водорода. Механизм переноса сводится к обратимому восстановлению пиридинового кольца в молекуле никотинамидных кофер-ментов, напр.:
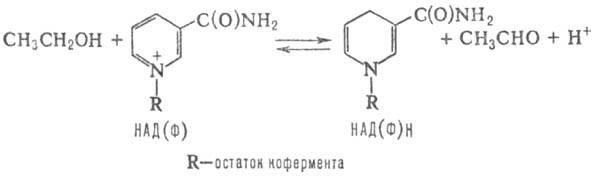
В результате таких р-ций НАД и НАДФ переходят в восстановленное состояние - НАДН и НАДФН. Когда последние выступают в качестве доноров электронов и протона, эти превращ. протекают в обратном направлении. Образование НАД(Ф)Н осуществляется стереоспецифично.
Функцион. различие между двумя коферментными формами состоит в том, что НАДН, как правило, служит поставщиком электронов в цепь биол. окисления, сопряженного с окислительным фосфорилированием, т. е. с процессами аккумуляции энергии в форме АТФ; НАДФН служит доно-ром водорода при биосинтезе жирных к-т и стеринов с участием ацетил-кофермента А; восстановит. аминировании кетокислот с образованием из них аминокислот (напр., глутаминовой к-ты из a-кстоглутаровой); восстановлении рибозы до дезоксирибозы; образовании восстановленных форм фолиевой к-ты (ди- и тетрагидрофолата; см. Фолацин); восстановлении глутатиона и метгсмоглобина; в разл. р-ц
…
Перейти к полному виду статьи
Свернуть
 В закладки будет добавлено толкование к данному слову в данном словаре. Закладки сохраняются на Вашем компьютере в cookie. Если Ваш браузер не поддерживает cookie или такая возможность отключена, то сохранение закладок будет не возможно.
В закладки будет добавлено толкование к данному слову в данном словаре. Закладки сохраняются на Вашем компьютере в cookie. Если Ваш браузер не поддерживает cookie или такая возможность отключена, то сохранение закладок будет не возможно.